Научный электронный журнал КубГАУ . № 06(14), 2005
УДК 581.450/8:582.734.3
ФОРМООБРАЗОВАНИЕ ЛИСТЬЕВ CONIUM MACULATUM L. В ТЕЧЕНИЕ ОНТОГЕНЕЗА
Наумов С. Ю. – к. с.-х. н., доцент
Луганский национальный аграрный университет
На основании проведенных исследований установлено, что разнообразие розеточных листьев болиголова пятнистого представляет собой последовательный ряд от тройчатосложных листьев к трижды-непарноперистосложным листьям. Листья цветоносного побега на протяжении онтогенеза проходят стадии от трижды-непарноперистосложных к тройчатосложным. Такое течение формогенеза листьев получило название Conium-типа.
Studies have shown that variety of bottom leaves of Conium maculatum presents itself consequent row from ternate to three-odd-pinnate compound leaves. Leaves of floriferous caulis pass stage from three-odd-pinnate compound to ternate during ontogeny. Such sequence of formation leaves has got a name Conium-type.
Conium maculatum L. (болиголов пятнистый, омег) – двулетнее травянистое растение, относящееся в подсемейству Apioideae Drude семейства Apiaceae Lindl. [1; 2], широко распространен на юго-востоке Украины [3], встречаясь как на лесных опушках, так и около жилья, у дорог, на свалках [1; 2; 4; 5], иногда как сорняк в посевах культурных растений, на пастбищах [6]. Все части растения ядовиты, особенно плоды, из-за наличия ядовитых алкалоидов [1; 2; 7]. Ранее препараты из C. maculatum широко применялись в медицине, но ввиду ненадежности лечебного действия и имевшихся случаев отравлений применение в качестве лекарства прекращено [1].
Изучение ботанической литературы показало, что, несмотря на широкую распространенность вида, сведения о морфологических особенностях C. maculatum скудны и в некоторых случаях противоречивы, в частности, это касается описаний строения листьев. Данные же об их строении являются важным признаком, использующимся в таксономии. К. Линней указывал, что для целей систематического описания растений лист "рассматривается в отношении простоты, сложности и размещения" [8]. К простым листьям он относил такие, у которых "черешок несет только один лист", сложные К. Линней описывал: "много листьев на одном черешке сообразно строению или степени сложности, которая определяется подразделением общего черешка". Например, трояко-перистые листья Линней определял так: "к черешку прикрепляются много двуперистых листочков" (рис. 1).

Рисунок 1 – Трижды-непарноперистосложный лист по К. Линнею [8]
Многие ботаники в прошлом столетии одним из основных признаков сложности листьев приняли их способность к раздельному опаданию листочков. В качестве примера сложного листа часто приводятся листья конского каштана [9]. Однако специально проведенные исследования опадания листьев Aesculus hippocastanum L. показали, что большая часть его листьев опадает целыми или с определенным количеством сохранившихся листочков в листе, свидетельствуя, таким образом, в пользу утверждения, что характер опадания листьев не имеет отношения к их подразделению на простые и сложные [10]. Способность к сбрасыванию листьев у растений является их приспособлением к переживанию неблагоприятных условий и для большинства травянистых растений не характерна. У растений данной жизненной формы, в том числе и у представителей семейства Apiaceae L., листья отмирают целиком, оставаясь при этом прикрепленными к стеблю, что не мешает их подразделению на простые и сложные.
В некоторых определителях растений листья C. maculatum описываются как черешковые, в очертании треугольные, трижды-перисторассеченные, с перисто-надрезанными продолговатыми долями; средние и верхние листья более мелкие и менее сложные, почти сидячие, с узким влагалищем [3; 5–7]. В других описаниях [1; 2; 4] дается более подробная характеристика долей и долек, снабженных черешочками, наличие которых характерно для сложных листьев, т.к. черешок и черешочки служат не только органами опоры и проведения, но и органами ориентировки листовых пластинок по отношению к солнцу [9]. Кроме того, листья болиголова пятнистого описываются как перисто-рассеченные, однако известно, что из долей состоят перисто-раздельные листья, перисто-рассеченные же расчленены на сегменты [6; 9]. Литературных описаний C. maculatum, в которых развитие его листьев рассматривалось бы с использованием онтогенетического подхода, найдено не было. Все вышеописанные неточности и пробелы и послужили побудительным мотивом для проведения специальных исследований.
Материалы и методы исследований
Семена Conium maculatum L. собраны во время ботанических экспедиций на территории Луганского природного заповедника. Наблюдения за развитием растений производили как в природных условиях, так и при выращивании их в лаборатории светокультуры при круглосуточном освещении, используя стандартные четырехламповые светильники дневного света промышленного производства. Освещенность в пределах 4000–7000 люкс.
Во время роста и развития растений проводили фенологические наблюдения, отмечая наступление каждой фазы развития растений. Каждый последующий полностью сформировавшийся лист обрывали с растения и закладывали в гербарную папку. После высыхания его изображение получали с использованием сканирующих устройств Epson GT 5000 и HP Scanjet 3770 (согласно разработанной методике [11]), а также цифровой фотокамеры Benq DC 3410.
Результаты исследований и их обсуждение
Conium maculatum, как и большинство представителей семейства Apiaceae L., на первом году вегетации формирует прикорневую розетку листьев. Семядольные листья болиголова являются первыми ассимилирующими органами проростка, они продолговатые, зеленого цвета, у исследуемых растений отмирали лишь после формирования шестого настоящего листа. Первый настоящий лист тройчатосложный длинночерешковый (рис. 2). Боковые листочки располагаются на коротких черешочках.
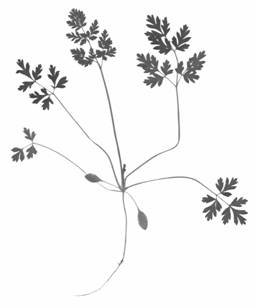
Рисунок 2 – Ювенильное растение Conium maculatum L.
Из рисунка видно, что все три листочка расчленены на три сегмента. Второй и третий листья морфологически сходны и в целом различаются размерами и глубиной расчленения. Они тройчатосложные, составляющие их листочки в свою очередь тройчато рассеченные. У третьего листа нижняя пара сегментов настолько обособлена от верхушечного сегмента, что их можно считать сидячими листочками. Верхушечные же сегменты разделены выемками на три доли. Верхняя доля также имеет тройчатую верхушку. Следующий, четвертый, лист следует отнести к дважды тройчатосложным, у которых в верхних частях листочков заложены потенции следующего листа, т.е. дальнейшее усложнение строения листьев связано с появлением дополнительных пар листочков, закладывающихся в акропетальной последовательности, т.е. чем выше на листе, тем моложе листочек. По этой причине верхние листочки менее крупные, менее дифференцированные и имеют черешочки меньшей длины, чем нижние. У C. maculatum через ряд промежуточных форм к концу первого года вегетации формируются крупные трижды-непарноперистосложные листья с продолговато яйцевидными непарноперисторассеченными листочками (рис. 3). По мере формирования новых листьев нижние листья постепенно отмирают, оставаясь на стебле в прикрепленном состоянии.
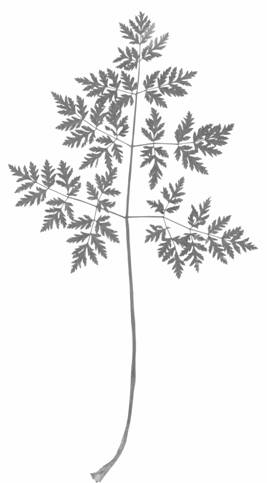
Рисунок 3 – Трижды-непарноперистосложный лист прикорневой розетки C. maculatum
На второй год после перезимовки первыми отрастают листья прикорневой розетки, которые по своему строению сходны с последними листьями первого года вегетации, т.е. они трижды-непарноперистосложные. В условиях достаточного увлажнения и высокой температуры эти листья могут достигать в длину 60 см и более, причем черешки у некоторых особей в средней части в диаметре превышают 2,5 см. Черешки округлые с едва заметными ребрами, количество которых обычно совпадает с количеством проводящих пучков в черешке. Следует отметить, что выше первого узла на рахисе с адаксиальной стороны появляется небольшой желобок, который от узла к узлу становится все глубже. Черешочки всех порядков ветвления желобчатые.

Рисунок 4 – Трижды-непарноперистосложный лист C. maculatum; сочленение черешка, рахиса с черешочками: а – срез нижней части листа с узлами 1-го и 2-го порядков; b – нижний узел (видны узлы 2-го и 3-го порядков); с – второй снизу узел с узлами 2-го и 3-го порядков; d – третий снизу узел; е – участок рахиса с верхними листочками
На рисунке 4а изображен участок листа, срезанный таким образом, что охватывает часть черешка, рахиса и узел второго порядка. Из рисунка видно, что черешок, рахис, а также черешочки имеют в разной степени выполненную центральную полость. Подчеркнем, что черешочки устроены принципиально так же, как черешки. В частности, крупные черешочки и первого, и второго порядков имеют в центре полость, что нехарактерно ни для листовой пластинки вообще, ни для ее центральной жилки. На рисунке 4b в увеличенном виде показан нижний узел сочленения черешка с черешочками первого порядка, причем черешок и черешочки располагаются явно не в одной плоскости, а под различными углами друг к другу, выполняя свойственные им функции расположения листовых пластинок наилучшим образом по отношению к свету. Такой же способ сочленения (ветвления) характерен и для узлов 2-го и 3-го порядков, а также и для вышележащих узлов на рахисе (рис. 4с и 4d). Подтверждением факта акропетального развития листа являются листочки верхушки листа, изображенные на рисунке 4е. Они меньше по размерам, менее расчленены, боковые листочки являются уже практически сидячими.
Во "Флоре СССР", как и в другой ботанической литературе, листья цветоносного побега описаны как "почти сидячие с узким влагалищем" [1; 2; 4]. На рисунке 5 представлен последовательный ряд листьев цветоноса C. maculatum.

Рисунок 5 – Последовательный ряд листьев цветоносного побега C. maculatum (последний лист увеличен в 8 раз)
Листья, изображенные на данном рисунке, уменьшены по сравнению с их реальными размерами, однако пропорции соблюдены. Из рисунка видно, что первые листья по строению не отличаются от развитых листьев прикорневой розетки. Они трижды-непарноперистосложные, длинночерешковые. Вышележащие листья претерпевают определенные изменения. От листа к листу уменьшаются линейные размеры, у них укорачивается и уплощается черешок, превращаясь в узкое влагалище. Через ряд промежуточных форм трижды-непарноперистосложные листья сменяются дважды-непарноперистосложными, а затем непарноперистосложными. В пазухах сидячих влагалищных листьев развиваются боковые цветоносные побеги, которые не закладываются в пазухах черешковых листьев. Наконец, последние листья главных цветоносов у значительного большинства исследованных растений болиголова тройчатосложные.
Таким образом, в онтогенезе Conium maculatum L. на первом году жизни при формировании листьев прикорневой розетки происходит их закономерное усложнение от тройчатосложных к трижды-непарноперистосложным. Последовательно формирующиеся листья цветоносного побега, наоборот, становятся все более простого строения. Основные этапы формогенеза листьев болиголова пятнистого представлены в виде схемы на рисунке 6. Это сделано в упрощенном виде без учета формы и расчленения листочков.
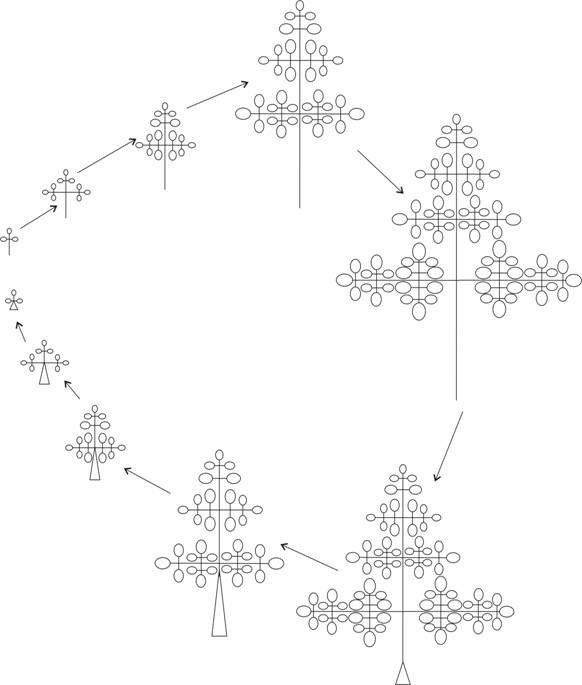
Рисунок 6 – Схема формообразования листьев C. maculatum в течение онтогенеза
Рассмотренный нами тип формогенеза листьев характерен для некоторых представителей семейства Apiaceae и, в частности, для Conioselinum tataricum Hoffm., Thyselium palustre (L.) Raf. и других. Назовем описанный в этой статье тип формообразования листьев Conium-типом. Два других типа, а именно Petroselinum-тип и Apium-тип установлены ранее [12; 13]. Описание других типов – задача последующих исследований.
Список литературы
1. Флора СССР / под ред. Б. К. Шишкина. – М.-Л. : Изд-во АН СССР. – 1950. – Т. XVI. – 648 с.
2. Флора УРСР / під ред. М. В. Клокова, О. Д. Вісюліна. – К. : Вид-во АН УРСР. – 1955. – Т. VII. – C. 461–618.
3. Мир растений. Краткий справочник-определитель / Авт.-сост. Д. Я. Зацепина, В. М. Остапко. – Донецк : Изд-во "Сталкер", 2002. – 304 с.
4. Ботанический атлас / под общ. ред. Б. К. Шишкина. – М.-Л. : Сельхозиздат, 1963. – 504 с.
5. Определитель высших растений Украины / Д. Н. Доброчаева [и др.]. – К. : Наукова думка, 1987. – 548 с.
6. Фисюнов, А. В. Определитель всходов сорняков / А. В. Фисюнов. – К. : Урожай, 1987. – 248 с.
7. Нейштадт, М. И. Определитель растений средней полосы Европейской части СССР / М. И. Нейштадт. – М. : Гос. уч.-пед. Изд-во Министерства просвещения РСФСР, 1963. – 640 с.
8. Линней, К. Философия ботаники / К. Линней. – М. : Наука, 1989. – 456 с.
9. Жуковский, П. М. Ботаника / П. М. Жуковский. – М. : Колос, 1982. – 623 с.
10. Наумова, И. А. Листопад у каштана конского (Aesculus hippocastanum L.) / И. А. Наумова // Збірник наукових праць Луганського НАУ. Серія Біологічні науки. – 2003. – № 22 (34). – С. 37–38.
11. Наумов, С. Ю. Применение компьютерной техники в биологии / С. Ю. Наумов, И. Д. Соколов, И. В. Миняева // Вісник ЛДПУ. – Луганськ : "Альма Матер", 2001. – № 6 (38). – С. 105–107.
12. Соколов, І. Д. Онтогенетичні закономірності формоутворення листя у Petroselinum crispum (Mill.) A. W. Hill. / І. Д.Соколов, С. Ю. Наумов, І. В. Міняєва, П. В. Шеліхов // Науковий вісник Волинського державного університету ім. Л. Українки. – Луцьк : РВВ "Вежа", 2002. – № 6. – С. 44–48.
13. ; Наумов, С. Ю. Apium-тип формообразования листьев в семействе Apiaceae L. / С. Ю. Наумов // Збірник наукових праць Луганського НАУ. Серія Біологічні науки. – Луганськ : Елтон-2, 2004. – № 39 (51). – С. 25–30.
Научный электронный журнал КубГАУ . № 06(14), 2005